Serie sobre nutrición ósea no desnaturalizada y salud ósea
I. Descripción general
Nuestros productos nutricionales no desnaturalizados para la salud ósea incluyen: Whole Bone Nutrition Powder , Non-Desnaturalized Bone Nutrition Ultra Micro Powder , Non-Desnaturalized Bone Protein Calcium Ultra Micro Powder y Non-Desnaturaled Type I Collagen Ultra Micro Powder . Estos productos están hechos de fémur de vaca fresco, apropiado para la edad (incluyendo articulaciones) como materia prima principal, lo que garantiza la más alta calidad de colágeno, glicosaminoglicanos y proteínas óseas. El proceso de fabricación implica ultramicroización a temperatura ultrabaja y tecnología de separación a temperatura ultrabaja basada en el principio de flujo de aire , lo que garantiza la preservación de la estructura no desnaturalizada . Esto preserva las propiedades funcionales de los ingredientes, haciéndolos ideales para apoyar la salud ósea y la reparación de tejidos.
La naturaleza única de estos productos, al no estar desnaturalizados, garantiza que conserven su estructura activa natural, esencial para que el cuerpo aproveche al máximo los compuestos bioactivos. Esta innovadora tecnología permite una absorción y eficacia superiores para favorecer la salud ósea y articular.
II. Relaciones de componentes

*El colágeno se compone principalmente de colágeno tipo I (más del 90 %), con una pequeña cantidad de colágeno tipo II (menos del 10 %).
III. Estructura activa natural
3.1 Estructura primaria
3.1.1 Colágeno tipo I: dominado por secuencias repetidas de aminoácidos Gly-Pro-X o Gly-X-Hyp (enlaces peptídicos)
3.1.2 Glucosaminoglicanos
Ácido hialurónico (AH): cadena de polisacárido lineal simple y repetitiva formada por enlaces glucosídicos β(1→3) y β(1→4) alternados entre el ácido D-glucurónico y la N-acetilglucosamina.
Sulfato de condroitina (CS): Una serie de cadenas de polisacáridos lineales microscópicamente heterogéneos con una estructura básica de ácido D-glucurónico y N-acetilgalactosamina unidos alternativamente, que presenta modificaciones de sulfatación complejas y variables en posiciones específicas de la N-acetilgalactosamina (y ocasionalmente del ácido glucurónico).
3.2 Estructuras superiores
3.2.1 Colágeno tipo I
Estructura secundaria: cadena peptídica α con estructura helicoidal levógira (principalmente enlaces de hidrógeno)
Estructura terciaria: Estructura de triple hélice (enlaces peptídicos, enlaces de hidrógeno, fuerzas de van der Waals, interacciones hidrofóbicas, enlaces cruzados covalentes de condensación aldólica)
3.2.2 Estructuras superiores de los glicosaminoglicanos (GAG)
Estructura superior del ácido hialurónico: forma una red enrollada aleatoriamente y altamente hidratada en solución.
Estructura superior del sulfato de condroitina: se une covalentemente a las proteínas centrales para formar proteoglicanos.
3.3 Superestructura (reticulación covalente supramolecular)
Entre macromoléculas de colágeno tipo I: estructura de reticulación de piridinolina.
Entre la osteocalcina y la hidroxiapatita (HAP): Calcio quelado por el ácido γ-carboxiglutámico.
IV. Mecanismo de acción y efectos
El polvo de nutrición ósea completa, el polvo ultrafino de nutrición ósea no desnaturalizada, el polvo ultrafino de calcio proteico óseo no desnaturalizado y el polvo ultrafino de colágeno tipo I no desnaturalizado no solo brindan nutrientes integrales y equilibrados esenciales para el crecimiento y la reparación del tejido óseo, sino que también brindan los beneficios de las "estructuras activas".
4.1 Mecanismo y efectos de la mejora de la absorción de calcio
4.1.1 Mecanismo: La estructura supercovalente (calcio quelado con ácido γ-carboxiglutámico) entre la osteocalcina presente en el polvo ultrafino de calcio de proteína ósea no desnaturalizada y la hidroxiapatita (HAP) está protegida por la estructura tridimensional de la proteína, lo que impide el contacto con el ácido gástrico. Esto permite que el complejo llegue intacto al intestino. Bajo la acción de las enzimas digestivas intestinales, se descompone en moléculas de calcio solubles, como el calcio quelado con ácido γ-carboxiglutámico o el calcio unido a péptidos de moléculas pequeñas. Esto aumenta la concentración de moléculas de calcio en el intestino, lo que permite su absorción a través de la vía paracelular impulsada por gradientes de concentración.
4.1.2 Efecto: Esta absorción depende de gradientes de concentración y es independiente de la saturación de la proteína transportadora de calcio (CaBP). En pacientes con niveles reducidos de CaBP debido al crecimiento, desarrollo o envejecimiento, lo que conlleva una absorción limitada de calcio, esto logra una compensación fisiológica de la captación de calcio.
4.2 Mecanismo y efecto de la promoción de la osteogénesis del calcio
4.2.1 Mecanismo: La osteogénesis del calcio está regulada por múltiples factores tanto en el entorno sanguíneo como en el del tejido óseo.
El entorno sanguíneo se ve influenciado principalmente por el coeficiente de concentración de calcio-fósforo. El polvo ultrafino de calcio de proteína ósea no desnaturalizada se digiere y descompone gradualmente en el intestino en moléculas de calcio solubles, como el calcio quelado con ácido γ-carboxiglutámico o el calcio unido a péptidos de moléculas pequeñas, lo que permite una absorción lenta. Su relación calcio-fósforo se acerca a 2:1, lo que maximiza la concentración de calcio-fósforo y promueve el flujo de calcio de la sangre al tejido óseo.
El entorno del tejido óseo está influenciado por tres factores: la nanorregión HAP del colágeno tipo I, la carboxilación de los residuos de ácido glutámico en la osteocalcina y la regulación mediante proteínas puente y moléculas de adhesión ósea.
La estructura de triple hélice de los fragmentos de colágeno tipo I no desnaturalizado que contiene puede activar el sistema linfático intestinal, regulando la inmunidad, suprimiendo las respuestas inflamatorias y ralentizando la degradación del colágeno tipo I. Sus productos de degradación (pequeñas moléculas como péptidos reticulados y péptidos conjugados) inducen la formación de la estructura del colágeno tipo I mediante múltiples mecanismos (sustrato, chaperona molecular, señalización). Al descomponerse en monómeros de aminoácidos, participan directamente en la síntesis de colágeno, proporcionando amplias nanoregiones para HAP y maximizando las nanoregiones colágeno-HAP.
Sus proteínas óseas no desnaturalizadas se unen al calcio para formar ácido γ-carboxiglutámico unido al calcio o pequeños péptidos unidos al calcio. Alternativamente, activan la γ-carboxiglutamil carboxilasa como sustratos o mediante transducción de señales, catalizando la carboxilación de tres residuos de ácido glutámico en la osteocalcina. Esto proporciona sitios de unión al calcio, maximizando los puntos de unión del calcio.
La desmina no desnaturalizada y la osteonectina no desnaturalizada que contiene pueden promover la síntesis de proteínas correspondientes, confinar espacialmente el HAP dentro del colágeno tipo I, anclar el colágeno al HAP y prevenir la pérdida de calcio óseo.
4.2.2 Efecto: El calcio puede entrar, almacenarse y permanecer.
4.3 Mecanismo y efectos del aumento del contenido de proteína ósea
Las proteínas óseas incluyen colágeno (principalmente colágeno tipo I) y glicoproteínas (principalmente osteocalcina, fibronectina, osteonectina y más de diez más). El mecanismo para aumentar su contenido es fundamentalmente similar, como lo demuestra el polvo ultrafino de colágeno tipo I no desnaturalizado.
4.3.1 Mecanismo
Mecanismo de regulación inmunitaria intestinal: El polvo ultrafino de colágeno tipo I no desnaturalizado posee una estructura tridimensional que resiste la degradación por los jugos gástricos, lo que permite el contacto físico directo con el sistema linfático intestinal. Esto activa las células T inmaduras para que se diferencien en Tregs específicos, suprimiendo así el ataque erróneo del sistema inmunitario al colágeno tipo I. Promueve la secreción de factores antiinflamatorios para inhibir la inflamación crónica y ralentizar la degradación del colágeno tipo I.
Mecanismo de regulación de la actividad enzimática intracelular: El polvo ultrafino de colágeno tipo I no desnaturalizado contiene estructuras de reticulación covalente primaria, alta y supercovalente intactas. Las enzimas digestivas intestinales descomponen selectivamente las reticulaciones covalentes primarias (enlace peptídico), preservando las altas y supercovalentes. Este proceso libera pequeñas moléculas, como péptidos reticulados y péptidos conjugados, o actúa como sustrato, chaperonas moleculares o moléculas de señalización para inducir la formación estructural del colágeno tipo I.
Aporte de nutrientes: Los péptidos reticulados, los péptidos conjugados y otras moléculas pequeñas se descomponen en aminoácidos individuales, participando directamente en la síntesis de colágeno.
4.3.2 Efectos
Esta regulación de doble acción (inhibir la degradación y promover la síntesis) produce una eficacia significativamente mayor que la función de suministro de nutrientes de las proteínas o péptidos desnaturalizados.
4.4 Mecanismo y efectos de la potenciación de los glicosaminoglicanos (GAG)
4.4.1 Mecanismo
Las partículas ultrafinas de glicosaminoglicano no desnaturalizadas liberan fragmentos de HA, CS y KS. Estos se digieren fácilmente en fragmentos de bajo peso molecular, oligosacáridos y unidades de disacáridos. Activan los condrocitos, inhiben las enzimas degradantes de la matriz (MMP o ADAMTS), regulan la inmunidad y ejercen efectos antiinflamatorios y analgésicos. Intracelularmente, se descomponen en moléculas individuales de glicosaminoglicano, participando directamente en la síntesis de HA, CS y KS.
4.4.2 Efectos
Aumenta el contenido de GAG.
V. Derivado de tejido
5.1 Derivado del tejido determinado por la estructura
Las funciones principales de la nutrición no desnaturalizada (inmunomodulación, regulación de la actividad enzimática) se logran mediante su estructura activa natural. Esta estructura presenta una alta especificidad, y determina las propiedades derivadas de los tejidos.
5.2 Naturaleza derivada del tejido de la serie de nutrición ósea no desnaturalizada
El polvo de nutrición ósea completa, el polvo ultrafino de nutrición ósea no desnaturalizada, el polvo ultrafino de calcio proteico óseo no desnaturalizado y el polvo ultrafino de colágeno tipo I no desnaturalizado utilizan huesos de pierna bovina frescos y apropiados para la edad (incluidas las articulaciones) como materia prima. Su proteína principal es el colágeno tipo I, y su naturaleza derivada de tejido debe determinarse con base en el colágeno tipo I. Si bien el colágeno tipo I del tejido cutáneo comparte homología genética con el tejido óseo y secuencias de aminoácidos idénticas, las diferencias en las modificaciones microambientales resultan en disposiciones distintivas de orden superior y ultraestructurales. Por lo tanto, el tejido cutáneo no se considera homólogo a la serie de nutrición ósea no desnaturalizada. Los tejidos homólogos primarios incluyen: tejido óseo duro, discos intervertebrales, meniscos y pequeñas cantidades de cartílago articular.
VI. Población idónea
6.1 Necesidades nutricionales: Apto para todas las personas con deficiencia de calcio, especialmente bebés, niños pequeños y ancianos.
6.2 Necesidades de crecimiento óseo: Adecuado para quienes requieren aumentar la altura durante las etapas de desarrollo y para quienes buscan retrasar la pérdida de altura durante el envejecimiento.
6.2 Necesidades de aumento de la masa ósea
6.2.1 Individuos con síntomas de raquitismo durante el crecimiento y desarrollo: tales como cierre retardado de la fontanela, fontanela cuadrada, cabeza plana, tórax en forma de paloma, tórax en embudo, cambios en forma de rosario, piernas arqueadas o rodillas valgas, escoliosis, etc.
6.2.2 Individuos de mediana edad y ancianos: Aquellos que presentan signos de hiperplasia ósea, pérdida de altura, cifosis, osteoporosis, hernias discales, etc.
6.3 Necesidades de reparación de la degeneración articular: Pacientes con osteoartritis degenerativa, especialmente aquellos con lesiones o inflamación de menisco.
6.4 Necesidades de curación de lesiones óseas: curación de lesiones óseas o crecimiento de implantes de cemento óseo, etc.
Nutrición articular no degenerativa - Serie sobre salud articular
I. Descripción general
Los productos nutricionales no desnaturalizados de la Serie Salud Articular incluyen: Nutrición Articular Total en Polvo, Nutrición Articular Ultrafina en Polvo No Desnaturalizada, Nutrición para Cartílago en Polvo No Desnaturalizada y Colágeno Tipo II Ultrafino en Polvo No Desnaturalizado. Todos son ingredientes alimenticios funcionales elaborados a partir de articulaciones de rodilla o cartílago bovino fresco, apropiado para la edad. Fabricados según las normas pertinentes mediante procesamiento ultrafino a temperatura ultrabaja (tecnología de invención nacional patentada) y tecnología de separación a temperatura ultrabaja mediante el principio de cabina de viento (tecnología propia).
II. Relaciones de componentes

*El colágeno se compone principalmente de colágeno tipo II (más del 90%), con una pequeña cantidad de colágeno tipo I (menos del 10%).
III. Estructura activa natural
3.1 Estructura primaria
3.1.1 Colágeno tipo II: compuesto principalmente de secuencias repetidas de aminoácidos Gly-Pro-X o Gly-X-Hyp (enlace peptídico)
3.1.2 Glucosaminoglicanos
Ácido hialurónico (AH): Una cadena de polisacárido lineal simple y repetitiva formada por enlaces glucosídicos β(1→3) y β(1→4) alternados entre el ácido D-glucurónico y la N-acetilglucosamina.
Sulfato de condroitina (CS): una serie de cadenas de polisacáridos lineales microscópicamente heterogéneas formadas por unidades alternadas de ácido D-glucurónico y N-acetilgalactosamina como estructura básica, con modificaciones de sulfatación complejas y variables que ocurren en posiciones específicas de las unidades de N-acetilgalactosamina (y ocasionalmente de ácido glucurónico).
3.2 Estructuras superiores
3.2.1 Colágeno tipo II
Estructura secundaria: cadena peptídica α con estructura helicoidal levógira (principalmente enlaces de hidrógeno)
Estructura terciaria: Estructura de triple hélice (enlaces peptídicos, enlaces de hidrógeno, fuerzas de van der Waals, interacciones hidrofóbicas, enlaces cruzados covalentes de condensación aldólica)
3.2.2 Estructura de orden superior de los glicosaminoglicanos (GAG)
Estructura de orden superior del ácido hialurónico: forma una estructura de red altamente hidratada y enrollada aleatoriamente en solución.
Sulfato de condroitina Estructura de orden superior: se une covalentemente a las proteínas centrales para formar proteoglicanos.
3.3 Estructura supramolecular (entrecruzamiento covalente supramolecular)
Reticulación intermolecular del colágeno tipo II: hidroxipiridolina.
Osteocalcina e hidroxiapatita (HAP): el ácido γ-carboxiglutámico quela el calcio.
IV. Mecanismo de acción y efectos
El polvo de nutrición para articulaciones completas, el polvo ultrafino de nutrición para articulaciones no desnaturalizadas, el polvo ultrafino de nutrición para cartílago no desnaturalizado y el polvo ultrafino de colágeno tipo II no desnaturalizado no solo brindan nutrientes integrales y equilibrados para los tejidos articulares, sino que también ejercen efectos "estructurales activos".
4.1 Mecanismo y efectos de la promoción de la absorción de calcio
4.1.1 Mecanismo: La estructura supercovalente (calcio quelado con ácido γ-carboxiglutámico) entre la osteocalcina y la hidroxiapatita (HAP) en el polvo ultrafino de calcio de proteína ósea no desnaturalizada evita el contacto con los jugos gástricos gracias a la protección de la estructura tridimensional de la proteína. Esto le permite llegar intacto al intestino, donde es descompuesto por las enzimas digestivas intestinales en moléculas de calcio solubles, como el calcio quelado con ácido γ-carboxiglutámico o el calcio unido a péptidos de moléculas pequeñas, lo que aumenta la concentración intestinal de calcio. Bajo el efecto del gradiente de concentración, el calcio se absorbe por la vía paracelular.
4.1.2 Efecto: Esta absorción depende de gradientes de concentración y es independiente de la saturación de la proteína transportadora de calcio (CaBP). En pacientes con niveles reducidos de CaBP debido al crecimiento, desarrollo o envejecimiento, lo que conlleva una absorción limitada de calcio, esto logra una compensación fisiológica de la captación de calcio.
4.2 Mecanismo y efecto de la promoción de la osteogénesis del calcio
4.2.1 Mecanismo: La osteogénesis del calcio está regulada por múltiples factores tanto en el entorno sanguíneo como en el del tejido óseo.
El entorno sanguíneo se ve influenciado principalmente por el coeficiente de concentración de calcio-fósforo. El polvo ultrafino de calcio de proteína ósea no desnaturalizada se digiere y descompone gradualmente en el intestino en moléculas de calcio solubles, como el calcio quelado con ácido γ-carboxiglutámico o el calcio unido a péptidos de moléculas pequeñas, lo que permite una absorción lenta. Su relación calcio-fósforo se acerca a 2:1, lo que maximiza la concentración de calcio-fósforo y promueve el flujo de calcio de la sangre al tejido óseo.
El entorno del tejido óseo está influenciado por tres factores: la nanoregión HAP del colágeno tipo I, la carboxilación de los residuos de ácido glutámico en la osteocalcina y la regulación por bridgina y osteonectina.
Los fragmentos de colágeno tipo I no desnaturalizado que contiene activan el sistema linfático intestinal, modulando la inmunidad, suprimiendo las respuestas inflamatorias y ralentizando la degradación del colágeno tipo I. Sus productos de degradación (pequeñas moléculas como péptidos reticulados y péptidos conjugados) funcionan como sustratos, chaperonas moleculares y vías de señalización para inducir la formación estructural del colágeno tipo I. La degradación posterior en monómeros de aminoácidos participa directamente en la síntesis de colágeno, proporcionando amplias nanoregiones para la HAP y maximizando las nanoregiones de HAP del colágeno.
Sus proteínas óseas no desnaturalizadas se unen al calcio para formar ácido γ-carboxiglutámico unido al calcio o pequeños péptidos conjugados con calcio. Alternativamente, activan la carboxilasa del ácido γ-carboxiglutámico como sustratos o mediante transducción de señales, catalizando la carboxilación de tres residuos de ácido glutámico en la osteocalcina. Esto proporciona sitios de unión al calcio, maximizando los puntos de unión del calcio.
La desmina no desnaturalizada y la osteonectina no desnaturalizada que contiene pueden promover la síntesis de proteínas correspondientes, confinar espacialmente el HAP dentro del colágeno tipo I, anclar el colágeno al HAP y prevenir la pérdida de calcio óseo.
4.2.2 Efecto: El calcio entra, se retiene y permanece.
4.3 Mecanismo y efectos de la reparación del tejido cartilaginoso
El cartílago articular es cartílago hialino, con colágeno tipo II como su proteína principal.
4.3.1 Mecanismo
Mecanismo de regulación inmunitaria intestinal: El polvo ultrafino de colágeno tipo II no desnaturalizado posee una estructura tridimensional que resiste la degradación de su conformación activa natural por los jugos gástricos. Llega al intestino e interactúa físicamente de forma específica con el sistema linfático intestinal, activando las células T vírgenes para que se transformen en Tregs específicos. Las Tregs modulan las células T efectoras para prevenir ataques inmunitarios erróneos al colágeno tipo II, secretan factores antiinflamatorios para suprimir la inflamación crónica y ralentizan la degradación del colágeno tipo II.
Mecanismo de regulación de la actividad enzimática intraorgánica: El polvo ultrafino de colágeno tipo II no desnaturalizado contiene estructuras de reticulación covalente primaria, de orden superior y de superorden intactas. Las enzimas digestivas intestinales degradan selectivamente las reticulaciones covalentes primarias (enlace peptídico), preservando las estructuras de orden superior y de superorden. Esto produce moléculas pequeñas, como péptidos reticulados y péptidos conjugados, o que sirven como sustratos, chaperonas moleculares o moléculas de señalización para inducir la formación de estructuras de colágeno tipo II.
Acción nutritiva: Las moléculas pequeñas como los péptidos reticulados y los péptidos conjugados se descomponen en aminoácidos que participan en la síntesis de colágeno tipo II.
4.3.2 Efectos
Esta regulación de doble acción (inhibir la degradación y promover la síntesis) produce una eficacia significativamente mayor que los efectos mediados por proteínas o péptidos desnaturalizados.
4.4 Mecanismo y efectos de los glicosaminoglicanos (GAG)
4.4.1 Mecanismo
Las partículas ultrafinas de GAG no desnaturalizadas proporcionan fragmentos de HA, CS y KS, que se digieren fácilmente en fragmentos de bajo peso molecular, oligosacáridos y unidades de disacáridos. Estos activan los condrocitos, inhiben las enzimas degradadoras de la matriz (MMP o ADAMTS), regulan la inmunidad y ejercen efectos antiinflamatorios y analgésicos. También pueden descomponerse intracelularmente en moléculas individuales de glicosaminoglicano, participando directamente en la síntesis de HA, CS y KS.
4.4.2 Efectos
Aumenta el contenido de GAG.
V. Derivado de tejido
5.1 Derivado del tejido determinado por la estructura
Las funciones principales de la nutrición no desnaturalizada (inmunomodulación, regulación de la actividad enzimática) se logran mediante su estructura activa natural. Esta estructura presenta una alta especificidad, y determina las propiedades derivadas de los tejidos.
5.2 Naturaleza derivada del tejido de la serie de nutrición articular no desnaturalizada
El Polvo Nutritivo para Articulaciones Completas, el Polvo Ultrafino de Nutrición Articular No Desnaturalizada, el Polvo Ultrafino de Nutrición para Cartílago No Desnaturalizada y el Polvo Ultrafino de Colágeno Tipo II No Desnaturalizado utilizan como materia prima articulaciones de rodilla bovinas frescas y apropiadas para la edad. Su proteína principal es el colágeno Tipo II, cuyo origen tisular se deriva de tejido cartilaginoso rico en colágeno Tipo II. Los tejidos homólogos primarios incluyen: cartílago articular y pequeñas cantidades de hueso duro.
VI. Población idónea
6.1 Necesidades de reparación de lesiones articulares
6.1.1 Necesidades de reparación de la osteoartritis degenerativa
6.1.2 Necesidades de reparación de traumatismos articulares
Nutrición cutánea no desnaturalizada - Serie Salud de la piel
I. Descripción general
Los productos nutricionales no desnaturalizados de la Serie Salud de la Piel incluyen: Polvo Nutritivo para la Piel Entera, Polvo Ultrafino Nutritivo para la Piel No Desnaturalizado y Polvo Ultrafino de Colágeno Tipo III No Desnaturalizado. Todos se elaboran a partir de pieles frescas de bovino o burro, apropiadas para su edad. Fabricados según las normas pertinentes mediante ultramicronización a temperatura ultrabaja (tecnología de invención nacional patentada) y tecnología de separación a temperatura ultrabaja mediante el principio de gabinete de viento (tecnología propia), estos ingredientes alimenticios funcionales no desnaturalizados.
II. Proporciones de ingredientes

III. Estructuras activas naturales
3.1 Estructura primaria
3.1.1 Colágeno tipo I/III: Principalmente secuencias de aminoácidos repetidas Gly-Pro-X o Gly-X-Hyp (enlaces peptídicos)
3.1.2 Glucosaminoglicanos
Ácido hialurónico (AH): cadena de polisacárido lineal simple y repetitiva formada por enlaces glucosídicos β(1→3) y β(1→4) alternados entre el ácido D-glucurónico y la N-acetilglucosamina.
Sulfato de condroitina (CS): una serie de cadenas de polisacáridos lineales microscópicamente heterogéneas formadas por unidades alternadas de ácido D-glucurónico y N-acetilgalactosamina como estructura básica, con modificaciones de sulfatación complejas y variables que ocurren en posiciones específicas de las unidades de N-acetilgalactosamina (y ocasionalmente de ácido glucurónico).
3.2 Estructuras superiores
3.2.1 Colágeno tipo I/III
Estructura secundaria: cadena peptídica α con estructura helicoidal levógira (principalmente enlaces de hidrógeno)
Estructura terciaria: Estructura de triple hélice (enlaces peptídicos, enlaces de hidrógeno, fuerzas de van der Waals, interacciones hidrofóbicas, enlaces cruzados covalentes de condensación aldólica)
3.2.2 Estructuras superiores de los glicosaminoglicanos (GAG)
Estructura superior del ácido hialurónico: forma una red altamente hidratada y enrollada aleatoriamente en solución.
Estructura superior del sulfato de condroitina: las proteínas centrales se unen covalentemente para formar proteoglicanos.
3.3 Superestructura (reticulación covalente supramolecular)
Reticulación intermolecular de colágeno tipo I: hidroxilisina piridinolina y lisina piridinolina.
IV. Mecanismo de acción y efectos
El polvo de nutrición para toda la piel, el polvo ultrafino de nutrición para la piel no desnaturalizada y el polvo ultrafino de colágeno tipo III no desnaturalizado no solo brindan nutrientes integrales y equilibrados esenciales para el crecimiento y la reparación del tejido de la piel, sino que también brindan los efectos de "estructuras activas".
4.1 Mecanismo y efectos de la mejora del colágeno
4.1.1 Mecanismo
Mecanismo de regulación inmunitaria intestinal: El polvo ultrafino de colágeno tipo I/III no desnaturalizado posee una estructura tridimensional que resiste la degradación de su estructura activa natural por los jugos gástricos. Llega al intestino e interactúa físicamente con el sistema linfático intestinal, activando las células T vírgenes para que se transformen en Tregs específicos (células T reguladoras). Las Tregs modulan las células T efectoras para prevenir ataques inmunitarios erróneos contra el colágeno tipo I/III no desnaturalizado, secretan factores antiinflamatorios para suprimir la inflamación crónica y ralentizan la degradación del colágeno tipo I/III no desnaturalizado.
Mecanismo de regulación de la actividad enzimática intracelular: El polvo ultrafino de colágeno I/III no desnaturalizado contiene estructuras de reticulación covalentes primarias, de alto orden y supercovalentes intactas. Las enzimas digestivas intestinales pueden seleccionar y descomponer específicamente las estructuras de reticulación covalentes primarias (enlace peptídico), preservando las estructuras de reticulación covalentes de alto orden y superorden. Este proceso produce pequeñas moléculas, como péptidos reticulados y péptidos conjugados, que inducen la formación de estructuras de colágeno de tipo I/III a través de múltiples vías, como sustratos, chaperonas moleculares o moléculas de señalización.
Función nutritiva: Las moléculas pequeñas como los péptidos reticulados y los péptidos conjugados pueden descomponerse en moléculas de aminoácidos individuales y participar directamente en la síntesis de colágeno.
4.1.2 Efecto
Esta regulación de doble acción (inhibir la degradación y promover la síntesis) produce una eficacia significativamente mayor que la función nutritiva proporcionada por las proteínas o péptidos desnaturalizados.
4.2 Mecanismo y efecto de la potenciación de los glicosaminoglicanos (GAG)
4.2.1 Mecanismo
Las partículas ultrafinas de glicosaminoglicano no desnaturalizadas proporcionan fragmentos de HA y CS que se digieren fácilmente en fragmentos de bajo peso molecular, oligosacáridos y unidades de disacáridos. Estas inhiben las enzimas que degradan la matriz (MMP o ADAMTS), regulan la respuesta inmunitaria para obtener efectos antiinflamatorios y analgésicos, y pueden descomponerse intracelularmente en moléculas individuales de glicosaminoglicano para participar directamente en la síntesis de HA y CS.
4.2.2 Efectos
Mejora el contenido de GAG.
V. Propiedades derivadas de tejidos
5.1 Propiedades derivadas del tejido determinadas por la estructura
Las funciones principales de la nutrición no desnaturalizada (inmunomodulación, regulación de la actividad enzimática) se logran mediante su estructura naturalmente activa. Esta estructura presenta una alta especificidad, y su composición define las propiedades derivadas de los tejidos.
5.2 Propiedades derivadas de los tejidos de la serie de nutrición de la piel no desnaturalizada
El Polvo Nutritivo para Piel Entera, el Polvo Ultrafino Nutritivo para Piel No Desnaturalizada y el Polvo Ultrafino de Colágeno Tipo III No Desnaturalizado utilizan como materia prima piel de burro o vaca fresca y apropiada para la edad. Sus proteínas principales son el colágeno Tipo I y III, y su origen tisular es el tejido cutáneo rico en estos tipos de colágeno. El principal tejido homólogo es el tejido cutáneo.
VI. Poblaciones adecuadas
6.1 Antienvejecimiento de la piel
6.2 Reparación de daños en la piel
Nutrición cerebral no desnaturalizada - Salud cerebral
I. Descripción general
El Polvo Ultrafino Neurotrófico de Cerebro No Desnaturalizado es un ingrediente alimentario funcional no desnaturalizado, producido a partir de cerebro fresco de bovino o porcino. Se fabrica de acuerdo con las normas pertinentes mediante molienda ultrafina a temperatura ultrabaja (tecnología de invención nacional patentada) y tecnología de separación a temperatura ultrabaja mediante el principio de cabina de viento (tecnología propia).
II. Relaciones de componentes
2.1 Clasificación general
Agua, lípidos, proteínas, sales inorgánicas y otras moléculas clave. Las sales inorgánicas y las moléculas clave están presentes en concentraciones muy bajas.

2.2 Composición de proteínas
Las proteínas del cerebro constituyen aproximadamente el 30% del peso seco y se pueden clasificar según su función:
2.2.1 Proteínas estructurales: proteínas de microtúbulos, microfilamentos (actina), proteínas de neurofilamentos.
2.2.2 Proteínas asociadas a sinapsis: proteínas de canales iónicos, proteínas receptoras de neurotransmisores, proteínas asociadas a vesículas, proteínas de andamiaje.
2.2.3 Proteínas asociadas a la mielina: proteína básica de la mielina, proteínas proteolipídicas, glicoproteínas asociadas a la mielina.
2.2.4 Metabolismo y proteínas relacionadas con la energía: Proteínas mitocondriales, hexoquinasa, enolasa
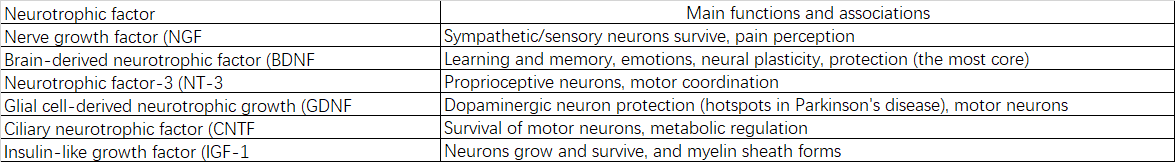
2.2.5 Factores neurotróficos:
2.2.6 Proteínas señalizadoras: proteínas G, proteína quinasas, fosfatasas, etc.
2.2.7 Proteínas asociadas a la enfermedad: β-amiloide, proteína tau, α-sinucleína.
2.3 Composición lipídica
2.3.1 Fosfolípidos: Cerebroside, fosfatidilinositol, fosfatidilserina, esfingomielina, lecitina, etc.
2.3.2 Colesterol
2.2.3 Grasas neutras
Dentro de los fosfolípidos y las grasas neutras, están presentes ácidos grasos específicos: los ácidos grasos saturados constituyen aproximadamente el 30% + , mientras que los ácidos grasos altamente insaturados representan alrededor del 60% + (ARA, DHA, ALA, EPA).
III. Relación entre las proteínas cerebrales y el envejecimiento
El envejecimiento cerebral no es un evento singular, sino un proceso sistémico y progresivo que implica el desequilibrio gradual de la red de homeostasis de proteínas del cerebro, el agotamiento gradual de proteínas funcionales y la acumulación gradual de proteínas anormales.
3.1 Declive integral de la red de homeostasis de proteínas
Los tres principales sistemas celulares que mantienen el control de calidad de las proteínas presentan un deterioro funcional:
3.1.1 Reducción de la eficiencia de la síntesis y plegamiento de proteínas
Las células envejecidas experimentan una disminución en las tasas de síntesis y fidelidad de proteínas, lo que aumenta la probabilidad de producir proteínas mal plegadas.
La respuesta de la chaperona molecular inducida por el estrés se torna menos efectiva y no logra ayudar adecuadamente a las proteínas recién sintetizadas a plegarse correctamente ni a reparar las proteínas dañadas.
3.1.2 Disminución de la capacidad del sistema de degradación de proteínas
El sistema ubiquitina-proteasoma exhibe una actividad reducida y no puede eliminar rápidamente las proteínas dañadas o mal plegadas; el ensamblaje del proteasoma también se ve afectado por el daño oxidativo, lo que acelera su declive funcional.
El sistema autofagia-lisosoma, en particular la macroautofagia y la autofagia mediada por chaperonas, muestra una eficiencia significativamente reducida. Esto impide la eliminación eficaz de agregados proteicos anormales y orgánulos dañados (p. ej., mitocondrias).
3.1.3 Deterioro del transporte y distribución de proteínas
La disfunción en las proteínas estructurales impide la entrega eficiente de las proteínas requeridas por las sinapsis y las mitocondrias al extremo distal, mientras que los productos de desecho no pueden transportarse rápidamente de regreso al cuerpo celular para su procesamiento.
3.2 Acumulación y toxicidad de proteínas anormales
El fallo de la red de homeostasis conduce a la acumulación gradual de diversas “proteínas de desecho” en el cerebro, dañando directamente las neuronas.
3.2.1 Acumulación de proteínas mal plegadas y propensas a la agregación
Aparecen oligómeros solubles de proteínas como β-amiloide, tau y α-sinucleína.
3.2.2 Acumulación de proteínas modificadas postraduccionalmente de forma anormal
Proteínas modificadas por AGE: los productos finales de la glicación avanzada se acumulan en el cerebro envejecido, lo que provoca reticulación de proteínas, endurecimiento, pérdida de función y activación de vías inflamatorias.
Proteína Tau hiperfosforilada: la Tau normal se desprende de los microtúbulos y pierde su función de estabilización del citoesqueleto.
3.3 Pérdida de proteínas funcionales clave
Además del aumento de “desperdicios”, las proteínas “útiles” también disminuyen o fallan.
3.3.1 Disminución de las proteínas asociadas a la sinapsis
La regulación negativa de proteínas presinápticas clave (por ejemplo, sinaptofisina, SNAP-25) y postsinápticas (por ejemplo, PSD-95, receptores NMDA/AMPA) causa directamente atrofia estructural sináptica y deterioro funcional, formando la base molecular directa para el deterioro del aprendizaje y la memoria.
3.3.2 Disminución de los factores neurotróficos y sus receptores
La síntesis y señalización debilitadas del factor neurotrófico derivado del cerebro (BDNF), el factor de crecimiento nervioso (NGF) y otros dan como resultado señales de apoyo insuficientes para la supervivencia, el crecimiento y la plasticidad neuronal.
3.3.3 Disminución de las proteínas reparadoras del ADN y de la actividad de la telomerasa
Esto conduce a la inestabilidad genómica y a la senescencia celular, lo que perjudica la supervivencia neuronal a largo plazo.
3.4 Consecuencias sistémicas
Las alteraciones a nivel de proteínas mencionadas anteriormente desencadenan una cascada de reacciones que culminan en el envejecimiento cerebral:
3.4.1 Disfunción mitocondrial: Las proteínas dañadas se acumulan en las mitocondrias, lo que reduce la producción de ATP y exacerba el estrés oxidativo.
3.4.2 Neuroinflamación crónica: Las células gliales (microglia y astrocitos) se activan de forma persistente por proteínas anormales (por ejemplo, oligómeros Aβ), transformándose de “guardianas” a “destructoras” al liberar factores proinflamatorios que dañan las neuronas sanas.
3.4.3 Pérdida de plasticidad sináptica: los desequilibrios en proteínas clave debilitan la potenciación a largo plazo, perjudicando la formación y consolidación de la memoria.
3.4.4 Reducción de la eficiencia de la red neuronal: el daño a las proteínas de mielina en la sustancia blanca y el transporte axonal deteriorado ralentizan la transmisión de información entre las regiones del cerebro y degradan la sincronización.
3.4.5 Desregulación de la autofagia y la apoptosis: en última instancia conduce a una pérdida neuronal selectiva, particularmente en regiones críticas como el hipocampo y la corteza prefrontal.
IV. Mecanismo de acción y efectos de la nutrición cerebral no degenerativa
La nutrición cerebral no degenerativa no solo proporciona nutrientes completos y equilibrados esenciales para el crecimiento y la reparación del tejido cerebral —incluyendo proteínas, fosfolípidos, AH, ALA, EPA, etc.—, sino que, aún más importante, aporta sus estructuras activas naturales. Estas estructuras poseen funciones especiales que van más allá de la mera nutrición.
4.1 Inhibición de la degradación funcional de proteínas cerebrales
Mecanismo: La estructura tridimensional del polvo ultrafino neurotrófico cerebral no desnaturalizado resiste la degradación por los jugos gástricos, lo que permite el contacto físico directo con los sistemas intestinal y linfático. Esto activa las células T inmaduras para transformarlas en Tregs (células T reguladoras) específicas. Las Tregs modulan las células T efectoras para prevenir ataques inmunitarios erróneos a las proteínas cerebrales, secretan factores antiinflamatorios para suprimir la inflamación crónica y ralentizan la degradación de las proteínas cerebrales.
4.2 Promoción de la formación estructural y reparación anormal de proteínas cerebrales
Mecanismo: El polvo neuroultrafino cerebral no desnaturalizado contiene estructuras de reticulación covalente primaria y secundaria intactas. Las enzimas digestivas intestinales descomponen selectivamente las reticulaciones covalentes primarias (enlace peptídico), preservando las secundarias. Este proceso produce pequeñas moléculas, como péptidos reticulados y péptidos conjugados, o sirve como sustrato para enzimas específicas que potencian la actividad catalítica; o funciona como chaperonas moleculares para reparar estructuras anormales; o induce la formación y reparación de estructuras anormales de proteínas cerebrales mediante múltiples vías, como la transducción de señales.
4.3 Suministro de nutrientes
Mecanismo: Las moléculas pequeñas como los péptidos reticulados y los péptidos conjugados pueden descomponerse aún más en aminoácidos individuales, participando directamente en la síntesis de proteínas cerebrales.
4.4 Efecto
Este doble efecto regulador de inhibir la degradación y promover la síntesis produce una eficacia significativamente mayor que la función de suministro de nutrientes de las proteínas o péptidos desnaturalizados.
V. Derivado de tejido
5.1 Regulación dependiente de la estructura derivada del tejido
Las funciones principales de la nutrición no desnaturalizada (inmunomodulación, regulación de la actividad) se logran mediante estructuras naturalmente activas. Estas estructuras presentan una alta especificidad y ejercen efectos reguladores únicamente sobre proteínas con estructuras idénticas.
5.2 Origen del tejido correspondiente
La nutrición cerebral no desnaturalizada utiliza tejido cerebral fresco bovino o porcino, incluyendo el cerebro, el mesencéfalo y el cerebelo. Por consiguiente, sus estructuras proteicas son homólogas a las presentes en el cerebro, el mesencéfalo y el cerebelo.
VI. Poblaciones adecuadas
6.1 Personas que requieren crecimiento y desarrollo cerebral: bebés, niños pequeños y aquellos que necesitan apoyo para el desarrollo cerebral y neuronal.
6.2 Personas que buscan tratamiento antienvejecimiento cerebral o tratamiento de disfunciones cerebrales.
Proteína muscular no desnaturalizada - Salud muscular
I. Descripción general
El polvo ultrafino de proteína muscular no desnaturalizada es un ingrediente alimentario funcional no desnaturalizado, elaborado a partir de solomillo de res fresco o pechuga de pollo. Se fabrica según las normas pertinentes mediante molienda ultrafina a temperatura ultrabaja (tecnología nacional patentada) y tecnología de separación a temperatura ultrabaja basada en el principio de cámara de aire (tecnología propia).
II. Razones y funciones de composición
2.1 Clasificación general

2.2 Composición y funciones de las proteínas
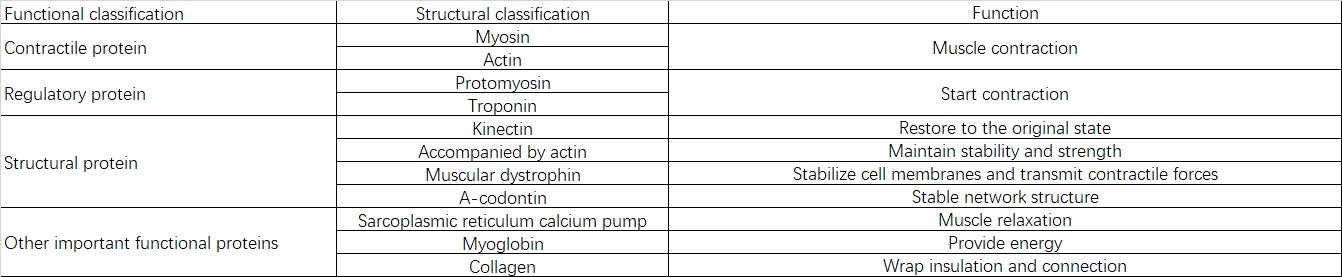
III. Causas de la sarcopenia
La sarcopenia se centra en la pérdida progresiva de masa, fuerza y función del músculo esquelético, resultado de múltiples factores interrelacionados.
3.1 Desequilibrio entre la síntesis y la degradación de proteínas (Patología central)
3.1.1 Síntesis reducida
Las tasas de síntesis de proteínas musculares disminuyen con la edad. Incluso bajo estimulación (p. ej., mediante nutrición o ejercicio), la respuesta sintética se debilita, lo que genera resistencia sintética.
3.1.2 Aumento de la degradación
Sobreactivación del sistema ubiquitina-proteasoma y del sistema autofagia-lisosomal, junto con la regulación positiva de genes como Atrogin-1 y MuRF1.
3.2 Desregulación de las vías hormonales y de señalización
3.2.1 Disminución de las hormonas anabólicas
Deterioro significativo del eje hormona del crecimiento/factor de crecimiento similar a la insulina-1.
Disminución de los niveles de hormonas sexuales (testosterona/estrógeno).
3.2.2 Inhibición de las vías anabólicas:
Resistencia a la insulina: reducción de la eficiencia de la captación y utilización de aminoácidos en el músculo.
Supresión de vías anabólicas clave: p. ej., disminución de la capacidad de activación de la vía de señalización mTOR.
Aumento de miostatina: inhibe el crecimiento muscular.
Señales catabólicas mejoradas: el aumento relativo o absoluto de los glucocorticoides promueve la descomposición de proteínas.
3.3 Inflamación crónica
El envejecimiento se acompaña de una inflamación sistémica, crónica y de bajo grado con citocinas proinflamatorias elevadas (por ejemplo, IL-6, TNF-α).
IV. Mecanismo de acción y efectos
El polvo ultrafino de proteína muscular no desnaturalizada no solo proporciona nutrientes integrales y equilibrados esenciales para el crecimiento muscular, sino que también posee propiedades “estructurales activas”.
4.1 Mecanismo y efectos de la inhibición de la degradación
Mecanismo: El micropolvo de proteína muscular no desnaturalizada contiene múltiples proteínas musculares no desnaturalizadas con estructuras tridimensionales intactas. Estas proteínas resisten la degradación por los jugos gástricos y llegan intactas al intestino delgado. Son fagocitadas por las células M, lo que permite un contacto físico específico con el sistema linfático intestinal. Esto activa las células T inmaduras para que se transformen en Tregs específicos. Esto modula los ataques erróneos de las células T efectoras a diversas proteínas musculares, secreta factores antiinflamatorios y suprime la inflamación crónica.
Efecto: Retarda la degradación de las proteínas musculares.
4.2 Mecanismo y efectos de la promoción de la síntesis
Mecanismo: El polvo ultrafino de proteína muscular no desnaturalizada contiene estructuras de enlaces cruzados covalentes primarios y de orden superior intactas. Las enzimas digestivas intestinales pueden seleccionar y descomponer específicamente las estructuras de enlaces cruzados covalentes primarios (enlace peptídico), preservando los enlaces cruzados covalentes de orden superior. Esto produce pequeñas moléculas, como péptidos reticulados y péptidos conjugados, que regulan la actividad catalítica de las proteasas de diversas maneras (como sustratos, chaperonas moleculares o moléculas de señalización) para mejorar la síntesis de proteínas musculares. Una mayor degradación en aminoácidos individuales permite la participación directa en la síntesis de proteínas musculares.
Efecto: Favorece la síntesis muscular.
Esta regulación de doble acción (inhibir la degradación y promover la síntesis) produce una eficacia significativamente mayor que la de las proteínas o péptidos desnaturalizados.
V. Derivado de tejido
5.1 Estructura derivada del tejido determinada
Las funciones principales de la nutrición no desnaturalizada (inmunomodulación, regulación de la actividad) se logran mediante estructuras naturalmente activas. Estas estructuras presentan una alta especificidad y ejercen efectos reguladores únicamente sobre proteínas con estructuras idénticas.
5.2 Origen específico del tejido
Los ingredientes de nutrición muscular no desnaturalizados provienen de tendón de res o pechuga de pollo, ambos tejidos musculares esqueléticos. El tendón de res presenta un 95 % de similitud estructural, mientras que la pechuga de pollo presenta un 85 % de similitud estructural.
VI. Poblaciones adecuadas
6.1 Atletas: Personas que realizan entrenamiento de fuerza y desarrollo muscular.
6.2 Sarcopenia: Inhibe la degradación muscular y promueve la síntesis muscular.
Proteína del bazo no desnaturalizada: salud inmunitaria
I. Descripción general
El polvo ultrafino de proteína de bazo no desnaturalizada es un ingrediente alimentario funcional no desnaturalizado, producido a partir de bazo bovino fresco. Se fabrica mediante tecnología de procesamiento ultrafino a temperatura ultrabaja (patente de invención nacional) y tecnología de separación a temperatura ultrabaja basada en el principio de cabina de viento (tecnología patentada), de conformidad con las normas pertinentes.
II. Relaciones y funciones de los componentes

2.1 Composición y funciones de los complejos de proteínas de respuesta inmunitaria central
2.1.1 Proteínas relacionadas con el sistema inmunitario (complejo funcional central): la fracción más distintiva y funcional dentro de las proteínas del bazo.
Composición: Anticuerpos (inmunoglobulinas IgG, IgM, etc.) y sus fragmentos.
Función: Ejecutores centrales de la inmunidad humoral.
2.1.2 Proteínas del complejo mayor de histocompatibilidad
Composición: Moléculas MHC de clase I y MHC de clase II, ampliamente expresadas en las superficies de las células presentadoras de antígenos.
Función: Capturar fragmentos de proteínas patógenas (antígenos) y “presentarlos” a los linfocitos T.
2.1.3 Proteínas del sistema del complemento
Composición: Múltiples componentes del complemento, incluidos C3, C4 y activadores del complemento.
Función: Formar el “complejo de ataque de membrana del complemento” para perforar directamente las membranas de los patógenos; actuar como “quimiocinas” para atraer células inmunes a los sitios de infección; marcar patógenos para promover la fagocitosis (actividad complementaria).
2.1.4 Citocinas y quimiocinas
Composición: Interleucinas, interferones, factores de necrosis tumoral, etc.
Función: Actúan como “moléculas de señalización” entre las células inmunes, dirigiendo con precisión el inicio, la mejora, la supresión o la terminación de las respuestas inmunes y coordinando el comportamiento de varios tipos de células inmunes.
2.1.5 Receptores de reconocimiento de patrones
Composición: Receptores tipo Toll, etc.
Función: Reconocer patrones moleculares conservados de patógenos para iniciar respuestas inmunes innatas rápidas.
2.2 Proteínas implicadas en el procesamiento de glóbulos rojos
2.2.1 Enzimas de degradación de la hemoglobina
Composición: Hemo oxigenasa, biliverdina reductasa, etc.
Función: Descompone la hemoglobina en los glóbulos rojos envejecidos, liberando iones de hierro para su reutilización y convirtiendo el hemo en bilirrubina.
2.2.2 Proteínas de almacenamiento y transporte de hierro
Composición: La ferritina es el representante más importante.
Función: Almacena y amortigua el hierro de forma segura y eficiente, evitando que el hierro libre genere radicales libres dañinos y libera hierro según lo necesite el cuerpo.
2.3 Proteínas estructurales y de andamiaje
Composición: Proteínas de la matriz extracelular (por ejemplo, colágeno, fibronectina) y moléculas de adhesión celular.
Función: Formar el andamiaje físico de la pulpa blanca del bazo (vainas linfáticas periarteriales, folículos linfoides) y la pulpa roja; mediar la localización de los linfocitos, la retención y las interacciones entre las células inmunes.
2.4 Metabolismo y enzimas antioxidantes
2.4.1 Enzimas antioxidantes
Composición: Superóxido dismutasa, catalasa, glutatión peroxidasa.
Función: Eliminación de especies reactivas de oxígeno (ROS) para proteger el tejido esplénico del daño oxidativo.
2.4.2 Enzimas metabólicas de nucleótidos
Composición: Nucleósido fosforilasa de purina, etc.
Función: Participa en el metabolismo de las purinas dentro de los linfocitos, fundamental para la función de las células inmunes.
2.5 Transducción de señales y proteínas reguladoras
Composición: Factores de transcripción que incluyen NF-κB, proteínas de la familia STAT, etc.
Función: Recibir señales como citocinas, translocarlas al núcleo para regular la expresión genética específica y determinar el destino de activación, proliferación o diferenciación de las células inmunes.
III. Mecanismo de acción y efectos
El polvo ultrafino de proteína de bazo no desnaturalizada proporciona proteínas de bazo intactas y con una conformación natural. Su mecanismo principal no implica el aporte de nutrientes, sino que funciona como un complejo depósito natural de moléculas bioactivas. Proporciona señales reguladoras y módulos funcionales al organismo, en particular al sistema inmunitario, ejerciendo así un efecto de "inmunidad nutricional".
3.1 Efectos inmunitarios intestinales locales
3.1.1 Efectos inmunes pasivos
Las partículas ultrafinas de proteína de bazo no desnaturalizada contienen abundantes inmunoglobulinas que llegan intactas al intestino bajo protección estructural. Sus fragmentos de unión a antígenos (regiones Fab) mantienen su actividad durante un tiempo, ejerciendo inmunidad pasiva local. Estos fragmentos Fab pueden: - Neutralizar directamente virus y bacterias específicos y sus toxinas, incapacitándolos para infectar o causar toxicidad; - Adherirse a patógenos para bloquear la invasión; - Formar complejos con patógenos o antígenos que estimulan la peristalsis intestinal y facilitan la encapsulación del moco, acelerando su expulsión a través de las heces y reduciendo su tiempo de retención y el daño intestinal.
3.1.2 Mantenimiento de la función de la barrera intestinal
Reducción del daño inflamatorio: Al eliminar rápidamente patógenos y toxinas, reduce los ataques directos a las células epiteliales intestinales y la respuesta inflamatoria excesiva resultante, protegiendo así la integridad de la barrera intestinal.
Apoyo a la reparación: algunos estudios sugieren que otros componentes de las preparaciones de inmunoglobulina (como los factores de crecimiento) o los propios anticuerpos pueden promover indirectamente la reparación de la mucosa.
3.1.3 Modulación del entorno inmunitario local
Amortiguación de antígenos: En el intestino, el órgano inmunitario más grande del cuerpo, el exceso de antígenos exógenos (incluidos los antígenos alimentarios y patógenos) puede desencadenar respuestas inmunitarias anormales. La inmunoglobulina oral se une y elimina una parte de estos antígenos, lo que reduce la carga sobre el sistema inmunitario intestinal y el riesgo de ataques mal dirigidos, contribuyendo así a mantener el equilibrio de la tolerancia inmunitaria.
Efectos antiinflamatorios: Al reducir la estimulación de patógenos a través de los mecanismos antes mencionados, regula negativamente la producción de citocinas proinflamatorias, creando un entorno local más propicio para la reparación de tejidos y el equilibrio microbiano.
3.2 Efectos inmunomoduladores sistémicos
3.2.1 Inducción de la tolerancia inmunitaria periférica como antígenos
El bazo contiene proteínas y antígenos relacionados con el sistema inmunitario en sus formas nativas, no desnaturalizadas. Estas sustancias pueden ser reconocidas por el sistema inmunitario intestinal como "estímulos inmunitarios amigables y familiares", lo que mejora la vigilancia y la capacidad de respuesta inmunitaria sin desencadenar una inflamación intensa (vía de tolerancia oral).
3.2.2 Suministro directo de componentes inmunológicamente activos y moléculas de señalización
Libera péptidos de moléculas pequeñas con actividad inmunomoduladora en el intestino. Estos fragmentos peptídicos funcionan como moléculas de señalización reconocidas por las células inmunitarias intestinales (p. ej., las de las placas de Peyer), estimulando y entrenando suavemente el sistema inmunitario, similar a un "ensayo inmunitario".
3.2.3 Interacción con el repositorio de información inmunitaria
Potencial para inducir tolerancia oral: Para ciertas predisposiciones autoinmunes, las teorías sugieren que la ingestión de tejidos que contienen antígenos autosimilares puede ayudar a inducir tolerancia inmunológica.
IV. Derivado de tejido
4.1 Estructura derivada del tejido dependiente
Las funciones principales de la nutrición no desnaturalizada (inmunomodulación, regulación de la actividad) se logran mediante estructuras naturalmente activas. Estas estructuras presentan una alta especificidad y ejercen efectos reguladores únicamente sobre proteínas con estructuras idénticas.
4.2 Origen del tejido correspondiente
El bazo, rico en diversas células y factores inmunes, corresponde al tejido inmune como su origen tisular.
V. Poblaciones adecuadas
5.1 Individuos inmunodeprimidos
5.2 Individuos con trastornos digestivos
Elastina no desnaturalizada: salud vascular
I. Descripción general
El polvo ultrafino de elastina no desnaturalizada es un ingrediente alimentario funcional no desnaturalizado, producido a partir de ligamentos nucales o aortas bovinas frescas. Se fabrica según las normas pertinentes mediante ultramicronización a temperatura ultrabaja (tecnología de invención nacional patentada) y tecnología de separación a temperatura ultrabaja basada en el principio de cabina de viento (tecnología propia).
II. Relaciones de componentes
2.1 Composición del polvo ultrafino de elastina no desnaturalizada (ligamento nucal bovino) y su polvo liofilizado

2.2 Composición del polvo ultrafino de elastina no desnaturalizada (aorta bovina) y su polvo liofilizado

III. Estructura activa natural
3.1 Estructura primaria
La secuencia de aminoácidos de la elastina consta de segmentos hidrofóbicos alternados (ricos en valina, alanina, glicina y prolina) y segmentos reticulados (ricos en lisina y alanina) dispuestos en tándem.
3.2 Estructura superior
La secuencia desordenada de aminoácidos de la proteína elastina impide la formación de estructuras secundarias regulares, al carecer de las estructuras convencionales de nivel II, III o IV. En su lugar, forma un polímero de red desordenada tridimensional mediante enlaces cruzados covalentes, como los de elastina.
IV. Mecanismo de acción y efectos
La elastina es una proteína no nutricional cuya función principal no es el aporte de aminoácidos, sino su “estructura activa natural” única, que ejerce efectos estructurales.
4.1 Mecanismo y efectos de la regulación inmunitaria periférica
4.1.1 Mecanismo: La estructura multidimensional exclusiva de la elastina no desnaturalizada activa la diferenciación de las células T linfoides de las placas de Peyer en células T reguladoras específicas (Treg). Esto modula el ataque erróneo del sistema inmunitario a la elastina endógena y suprime las respuestas inflamatorias.
4.1.2 Efectos: Retarda la degradación de la elastina. Con el envejecimiento (que comienza alrededor de los 25-30 años), la capacidad del cuerpo para sintetizar elastina disminuye drásticamente. Simultáneamente, debido a la desregulación inmunitaria, las fibras de elastina existentes se fracturan, endurecen y dejan de funcionar debido a la glicación, el estrés oxidativo (daño por radicales libres) y la degradación enzimática (p. ej., por metaloproteinasas de matriz). Esta es la principal causa del endurecimiento vascular en el cuerpo humano.
4.2 Como molécula de señalización celular (función más destacada)
4.2.1 Mecanismo: La superficie de las moléculas de elastina intactas contiene dominios bioactivos específicos (p. ej., dominios de unión celular, dominios quimiotácticos). Cuando estos dominios mantienen su conformación nativa, pueden unirse específicamente a receptores (p. ej., proteínas de unión a elastina) en las superficies de fibroblastos, células endoteliales y otras células. Esta unión indica a las células que estimulen su propia síntesis de colágeno, elastina y ácido hialurónico adicionales. Este proceso induce la regeneración endógena, que es mucho más importante que el simple suministro de materias primas.
4.2.2 Efecto: Contribuye a mejorar fundamentalmente la elasticidad y firmeza, restaurando la elasticidad de tejidos como los vasos sanguíneos.
4.2 Como sustrato modulador de la actividad enzimática
4.2.1 Mecanismo: La elastina no desnaturalizada puede descomponerse en el intestino en péptidos reticulados que contienen estructuras de enlaces cruzados covalentes primarios y secundarios. Estos péptidos reticulados sirven como sustratos para enzimas de modificación estructural relevantes, lo que regula positivamente su actividad catalítica.
4.2.2 Efecto: Promueve la formación o reparación de estructuras de elastina.
4.3 Como plantilla estructural y andamio
4.3.1 Mecanismo: La elastina no desnaturalizada, que retiene grandes fragmentos estructurales y estructuras reticuladas no desnaturalizadas, puede funcionar como un "andamio guía" in vitro o in vivo. Dirige las moléculas precursoras de elastina recién sintetizadas hacia una disposición ordenada y una correcta reticulación, facilitando así la formación de nuevas redes funcionales de fibras elásticas.
4.3.2 Efecto: Ayuda a reparar fibras elásticas rotas y desordenadas causadas por el fotoenvejecimiento o el envejecimiento.
V. Derivado de tejido
5.1 Estructura derivada del tejido dependiente
Las funciones de las proteínas no desnaturalizadas (inmunomodulación, regulación de la actividad) dependen de la estructura. Su estructura presenta una alta especificidad, regulando únicamente proteínas con estructuras idénticas.
5.2 Fuentes derivadas de tejidos correspondientes
Tejidos ricos en elastina, como los vasos sanguíneos, la piel y los alvéolos.
VI. Candidatos idóneos
6.1 Salud vascular: hipertensión, hiperlipidemia, aterosclerosis.
6.2 Antiarrugas de la piel: Flacidez de la piel, arrugas, pérdida de elasticidad.
6.3 Insuficiencia Pulmonar: Pacientes con enfisema, enfermedad pulmonar obstructiva crónica (EPOC), etc.





